Технология криоконсервации спермы, несмотря на большое разнообразие криопротекторов,
не является совершенной и процесс криоконсервации приводит к потере
примерно 50% жизнеспособности сперматозоидов [1]:
Криоконсервация спермы предназначена не только для поддержания
первоначальной подвижности сперматозоидов, но и для сохранения необходимого
обмена веществ, что, однако, в процессе замораживания-оттаивания клеток не
всегда достигается в силу криогенного повреждения белков, липидов и
нуклеиновых кислот [2, 3, 4]. При криоконсервации увеличиваются окислительные процессы, растет
уровень свободных радикалов, обладающих высокой реакционной способностью к
повреждению клеточных и субклеточных структур, снижается жизнеспособность
клеток [5]. Поскольку
окислительные процессы являются триггерами морфологических и биохимических
криогенных повреждений, вызывающих дисфункцию сперматозоидов, повреждение клеточных
мембран, снижение активности ферментов, целесообразность использования
антиоксидантов, позволяющих ограничить действие процесса окисления, является
перспективным подходом. Использование молекулярного водорода (H2) в
качестве универсального антиоксиданта сегодня широко обсуждается и
рассматривается его терапевтическое действие, основанное на антиоксидантных
свойствах, в различных областях медицины [6, 7]. При этом
обоснование эффективности использования H2 в криоконсервации на
сегодняшний день в современной литературе не представлено. Объективная оценка
действия H2 возможна только при проведении комплексного анализа состояния
сперматозоидов. Новые перспективы в изучении клеточных процессов появились в
связи с разработкой интерференционных методов оптической микроскопии [8]. Визуализация
структурных и морфологических изменений клеток с высоким пространственным
разрешением является информативным способом изучения их уникальных
физиологических и биомеханических свойств [9]. Использование интерференционной
лазерной микроскопии и расшифровка интерференционных изображений дает
возможность оценить прозрачные биообъекты, без дополнительного применения
красителей, с анализом плотности внутриклеточных структур [10].
Целью работы ставилось изучение изменения морфо-функционального
состояния сперматозоидов в процессе криоконсервации при действии H2
с использованием лазерной интерференционной микроскопии.
Морфологические параметры и результирующие характеристики, полученные методом интерференционной микроскопии, основаны на анализе показателя преломления объекта и его морфологии. Результатом интерференционной микроскопии является не просто отражение деталей поверхности объекта, а регистрация оптической плотности объекта прямым и отраженным пучком света, что определяет принципиальное отличие результатов, полученных путем обычной световой микроскопии и интерференционной лазерной микроскопии (рис. 1). Данная особенность позволяет проводить анализ морфологии клеток в зависимости от плотности внутриклеточных структур клеток для изучения их физиологии.

Рис. 1. Автоматизированный интерференционный микроскоп. 1 – автоматизированный двухкоординатный предметный стол; 2 – опорное зеркало на пьезоэлементе; 3,5 – ПЗС-камеры; 4 – микроинтерферометр; 6 – лазерный осветитель.
Появление конгломератов и плотных структур определяет увеличение коэффициента пpеломления и регистрацию выпуклых доменов, для чего используется наклонное освещение с разных ракурсов, достигаемое за счет смещения точечного источника в плоскости апертурной диафрагмы микроскопа Реконструкция фазы проводится метод фазовых шагов [11]. При этом на фотоприемнике проводится регистрация смещения исследуемого объектного пучка относительно опорного пучка с известной фазой, что представляет интерферограмму объекта (рис. 2).
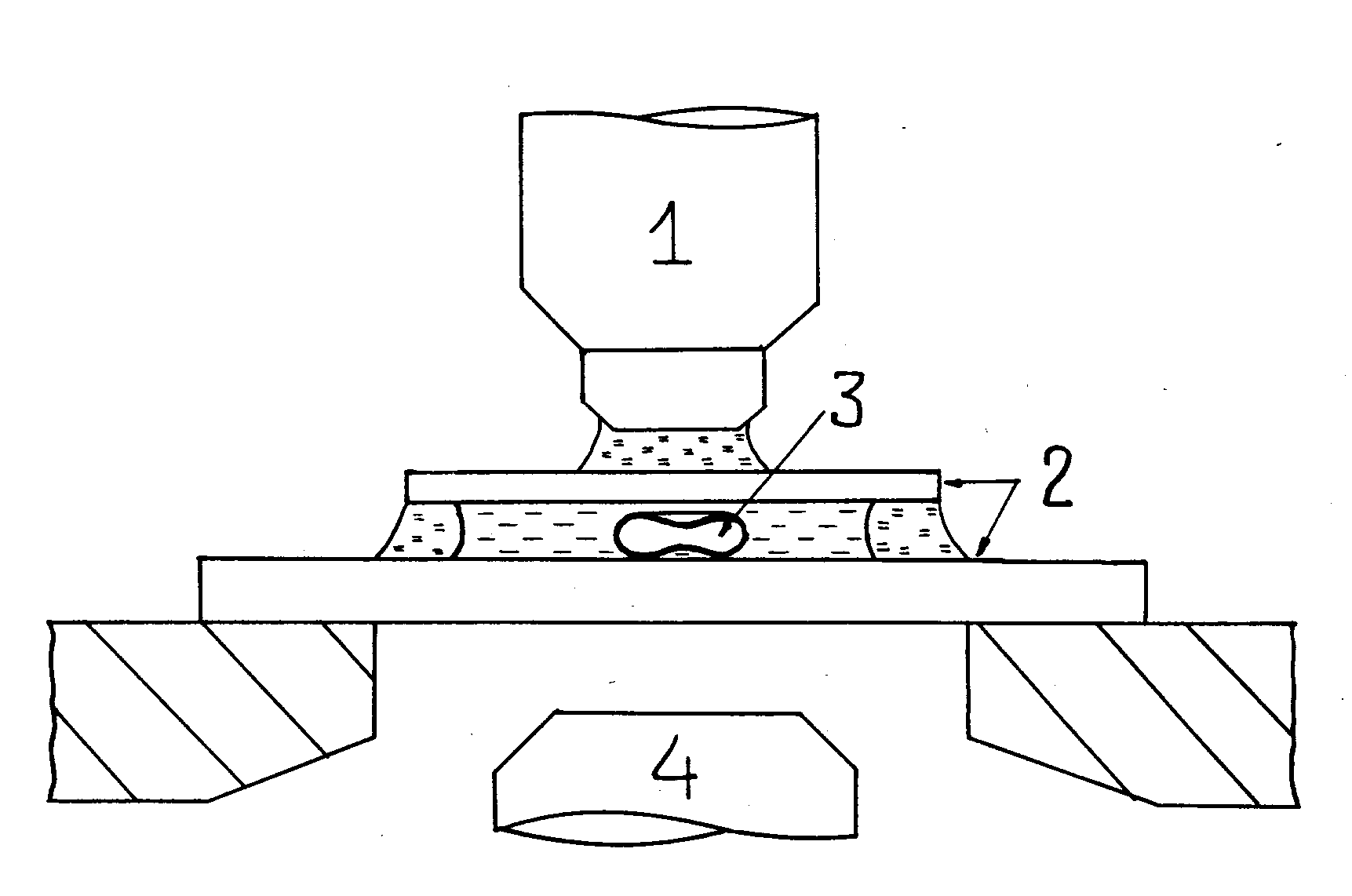
Рис. 2. Схематическое изображение микрокамеры и (в увеличенном масштабе) фотометрируемой клетки. 1 – объектив микроскопа, 2 - покровные стёкла, 3 - эритроцит, 4 – конденсор.
Расчет пространственного
распределения интенсивности получаемого объекта в плоскости проводится по
следующей формуле:
|
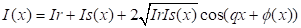
|
(1)
|
Где Ir и Is интенсивности в опорном и объектном плечах
интерферометра, q - пространственная частота полос,
φ - фаза, связанная с объектом.
Для прозрачных объектов
Is(x)
имеет слабую зависимость от x.
При помощи настройки увеличения системы можно выбрать частоту
q близкую или превышающую максимальную частоту
интерференционных полос, ограниченную числовой апертурой объектива, поэтому
фундаментальное дифракционное разрешение разрешающей способности сохраняется.
Интерференционный член
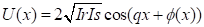 может быть выделен при помощи
высокочастотной Фурье фильтрации. Из этого следует, что комплексный
аналитический сигнал, связанный с действительной функцией u(x) может быть получен как
может быть выделен при помощи
высокочастотной Фурье фильтрации. Из этого следует, что комплексный
аналитический сигнал, связанный с действительной функцией u(x) может быть получен как
|
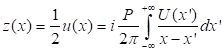
|
(2)
|
В равенстве (2) с
использованием преобразование Гильберта для u(x), фаза, связанная с комплексным аналитическим сигналом
z может
быть представлена следующим образом:
|
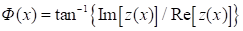
|
(3)
|
Базовой основой
интерференционного микроскопа является модифицированный интерферометр
Маха-Цендера. Источником излучения в интерферометре Маха-Цендера является
He-Ne лазер [8]. Опорное поле для создания
интерференционного изображения наклонено по отношению к объектному пучку под углом
45° по отношению к осям x и y
Схема, используемого в данной работе
лазерного интерференционного микроскопа, приведена на рис. 3. Прибор включает источник
света I, полупрозрачные зеркала BS1, BS2, BS3, поворотные зеркала M1, M2,
поляризаторы WP1, WP2, фазовый модулятор PM, объектива O, анализатора A и
детектора D. Поляризаторы WP1, WP2 модулируют фазы опорного и объектого пучков.
Фазовый модулятор PM формирует модуляцию фазы опорного пучка. Объектный пучок отражается
от объекта исследования, находящегося на столике S, проходит через зеркало BS3,
смешивается с опорным пучком и проходит через анализатор А, который выделяет из
него компоненту с той или иной поляризацией, и попадает на детектор.
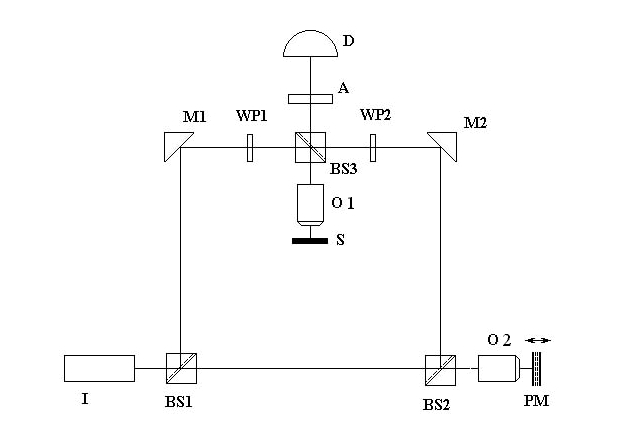
Рис. 3. Схема интерферометра, лежащая в
основе интерференционного лазерного микроскопа.
Использование
интерференционного лазерного микроскопа позволяет достигать сверхвысокого
разрешения, которое достигает 0,1 нм (по вертикали) и 15 - 100 нм в плоскости
объекта [12].
Расчет интерференционной
картины, полученной из модели объекта, позволяет представить выражение
трехмерной формы клеток, является новым подходом к визуализации для анализа
морфологии клеток, который может быть полезен для изучения их уникальных
физиологических и биомеханических свойств.
В работе исследовали сперму, разбавленную средой «BioXcell»
(Франция), затем проводили ее заморозку в жидком азоте (-196°С) по ГОСТу 26030-2015. Анализировали
состояние сперматозоидов после размораживания спермы по
стандартной технологии. Исследовали раз веденную разбавителем «BioXcell» сперму (группа I),
сперму после глубокой заморозки (группа II) и
сперму после глубокой заморозки, предварительно обработанную
Н2 (группа III). Концентрация H2 в
растворе находилась в пределах 1,2-1,5 мг/л.
Барбитурирование среды H2
производили с использованием генератора
молекулярного водорода «Спутник-3» (Китай).
Для оценки качественных показателей сперматозоидов использовали
спермоанализаторы «Биола АФС-500» НПФ БИОЛА (Россия). Энергетические показатели
сперматозоидов оценивали по концентраци и АТФ неэнзиматическим методом [13]. Антиоксидантную систему
анализировали по активности СОД [14] и каталазы [15]. Окислительные свойства клеток
определяли по концентраци и МДА в сперматозоидах по реакции
с тиобарбитуровой кислотой [16].
Интерференционную
лазерную микроскопию проводили на микроскопе
МИМ-340 с использованием лазера с длиной волны 650 нм [17]. Разрешение
по вертикали составляло 0,1 нм. Интерфореграммы обрабатывали в
программе MIM Visualizer 1.0 (MIM Software Inc., США).
Различия
между группами сравнивались с использованием критерия Стьюдента
с поправкой Бонферрони с учетом порога значимости p≤0.05.
Корреляционный анализ проводили с использованием коэффициента корреляции Спирмена.
Расчетные показатели интерфореграмм
сперматозоидов без воздействия (I группа) составили:
фазовая высота 24,03±0,02 нм, длина головки и хвоста
сперматозоидов 9,53±0,62 мкм и 46,82±5,25 мкм соответственно (табл. 1). Криоконсервация
определила уменьшение длины головки и хвоста у 7,01% и у 9,53% сперматозоидов
соответственно (р≤0,05)
.
Использование H2
в процессе криоконсервации
привело к сохранению оптико-геометрических параметров сперматозоидов на уровне нативных
клеток (I группа).
Таблица 1. Влияние криоконсервации и молекулярного
водорода на оптико-геометрические параметры сперматозоидов
|
Показатели сперматозоидов
|
Нативные
сперматозоиды, (группа I)
|
Сперматозоиды после
криоконсервации, (группа II)
|
Сперматозоиды после
воздействия молекулярным водородом и криоконсервации, (группа III)
|
|
Длина головки, мкм
|
9,53±0,62
|
8,32±0,55*
|
9,04 ±0,49
∆
|
|
Длина хвоста, мкм
|
46,82±2,05
|
42,36±2,35*
|
45,74±1,59 ∆
|
|
Фазовая высота, нм
|
24,03±0,02
|
21,32±0,05*
|
23,04 ±0,04*, ∆
|
Примечание:
* – различия по отношению к группе I, р≤0,05;∆ – различия между группами
после криоконсервации (группа II и группа III).
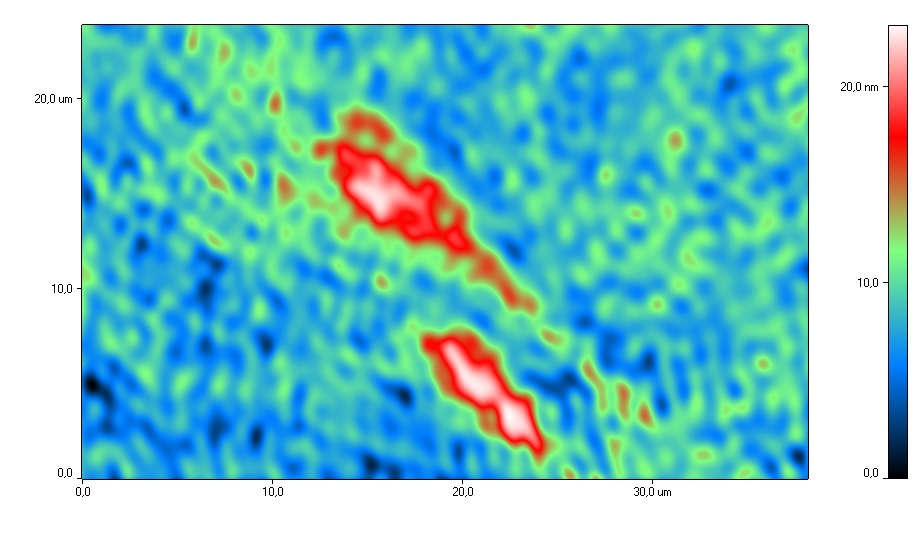
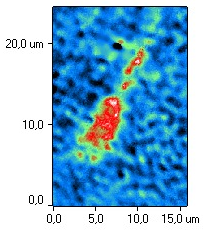
На рис. 4 представлены типичные фазовые портреты
сперматозоидов при действии H2 в ходе криоконсервации и без H2 при криоконсервации.
Анализ фазово-интерференционных изображений сперматозоидов
показал, что после воздействия Н2, головка сперматозоида была овальной
формы, средняя часть клетки была тонкой, хвост – прямой. Акросомальная область
после воздействия молекулярным водородом была хорошо
определяемой и занимала от 40 до 70% клетки. Такое распределение акросомы
соответствует физиологической норме, поскольку показано, что акросома покрывает около 2/3 передней
поверхности головки. Определение статуса акросомы в криоконсервированной сперме
имеет основополагающее значение, так как криоконсервация непосредственно наносит
ущерб мембранам сперматозоидов, что может привести к потере содержимого
акросомального матрикса. После криоконсервации без Н2 отмечено неравномерное распределение цитоплазмы в области
головки с аномальной акросомой, свидетельствующее
об изменении проницаемости плазматической мембраны и потере способности прикрепляться к мембране
ооцита. Кроме того, отмечается, что потеря акросомального содержимого матрицы
снижает долговечность криоконсервированных сперматозоидов [18].
|
|
|
|
а)
|
б)
|
Рис. 4. Типичные фазовые изображения сперматозоидов после криоконсервации
с использованием H2 (а) и после криоконсервации без H2 (б).
Оценка фазовых высот интерфореграмм
сперматозоидов и изменений показателя преломления позволила оценить
распределение плотности вещества в клетках. Фазовая высота в плоскости
представляет собой пространственную модуляцию волны от когерентного источника, которая
преобразуется в двумерное распределение
оптической разности хода и
определяется клеточным содержимым, обладающим различными показателями
преломления в различных компартаментах клетки [19].
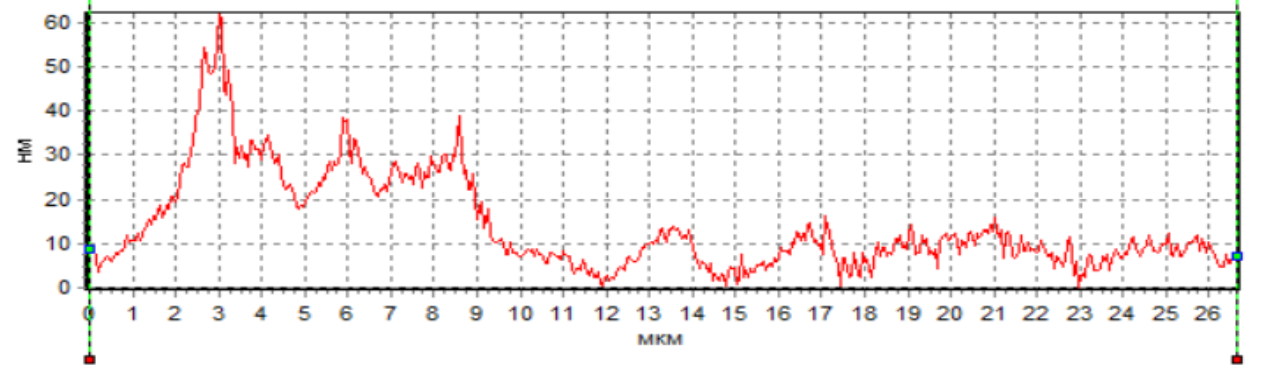
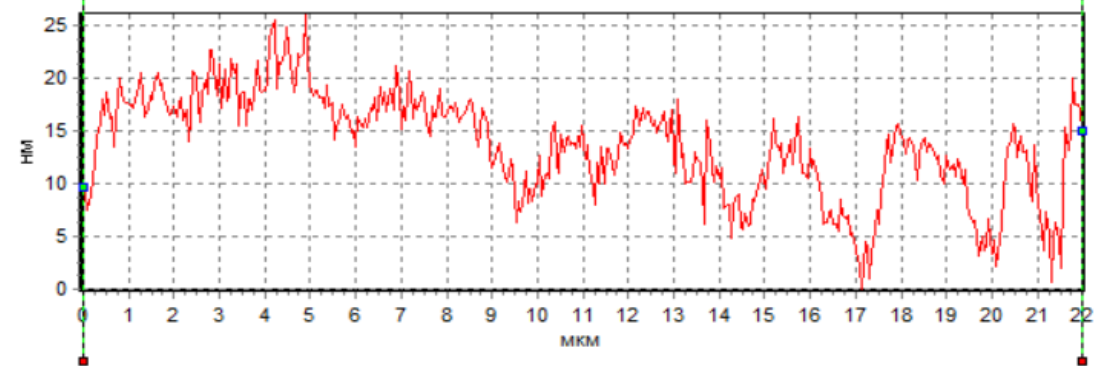
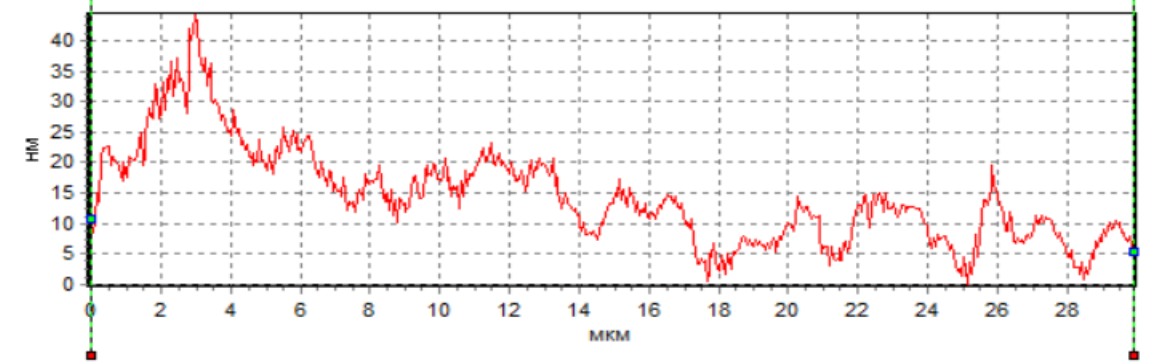
Анализ профиля сперматозоидов показал наличие фазового пика в области начального сегмента головки в нативных сперматозоида и при действии H2 в ходе криоконсервации (рис. 5а, 5в), что cвидетельcтвует о макcимальной плотноcти в акросомальной области. Кроме того, следует отметить, что как в нативных смерматозоидах, так и при использовании H2 в ходе криоконсервации фазовая высота в области акросомы значительно превосходила фазовую высоту у сперматозоидов при криоконсервации без H2 (рис. 5). Анализируя результаты, необходимо учитывать, что данная область соответствует ядру с ядрышком. Однако, уменьшение фазовой высоты сперматозоидов при криоконсервации без H2 свидетельствует об уменьшении плотности в этой области клетки, что может быть обусловлено потерей белка. В частности показано, что изменение в белковом составе мембраны сперматозоидов является одной из основных причин снижения фертильности сперматозоидов после криоконсервации, поскольку белки участвуют в процессах, которые носят основополагающий характер для оплодотворения: подвижность, реакция акросомы, слияние с яйцеклеткой [20].
Рис. 5. Пpофили изменения фазовой выcоты сперматозоидов: интактных (а), после криоконсервации без использования H2 (б) и после криоконсервации с использованием H2 (в).
Данное положение подтверждают ранее выявленные факты, полученные с использованием интерференционной микроскопии, свидетельствующие, что изменение фазовой высоты пpопоpционально пpоизведению толщины мембpаны на pазноcть локального показателя пpеломления клетки и pаcтвоpа [21].
В наших экспериментах при действии криоконсервации наблюдалось снижение фазовой высоты сперматозоидов (рис. 5б). По всей видимости, это связано с повреждением сперматозоидов в ходе криоконсервации и последующим их оттаиванием. В частности отмечено, что фазовые высоты уменьшаются пропорционально степени повреждения и увеличения количества воды внутри клетки, что приводит к уменьшению показателя преломления вследствие потери концентрации вещества внутри [22]. Разрушение плазматической мембраны во время кроиокнсервации может вызвать дальнейшее повреждение клеток и, следовательно, привести к необратимому повреждению их целостности. Во многом данное развитие событий может быть обусловлено окислительными процессами и изменением метаболизма клеток, поскольку интерфореграммы отражают динамику различных внутриклеточных процессов [23]. Для обоснования высказанного положения нами были проведены параллельные с интерференционной регистрацией клеток биохимические исследования внутриклеточного метаболизма.
Исследование уровня окислительного метаболизма выявило, что при криоконсервации регистрировалось значительное увеличение процесса перекисного окисления липидов (ПОЛ), что характеризовалось ростом концентрации малонового диальдегида (МДА) на 39% относительно нативных сперматозоидов (группа I) (табл. 2). Использование H2 в процессе криоконсервации позволило сохранить интенсивность окислительных процессов на уровне нативных сперматозоидов.
Таблица 2. Окислительная
и метаболическая активность сперматозоидов при криоконсервации и действии
молекулярного водорода
|
Показатели
|
Нативные
сперматозоиды, (группа I)
|
Сперматозоиды после
криоконсервации, (группа II)
|
Сперматозоиды после
воздействия молекулярным водородом и криоконсервации, (группа III)
|
|
МДА, нМоль/мл
|
0,61±0,12
|
0,85±0,14*
|
0,56±0,08 ∆
|
|
СОД, ед.акт./мг белка
|
0,61±0,08
|
0,78±0,08*
|
0,87±0,08*, ∆
|
|
Каталаза, мкат/мг
|
9,03±0,76
|
8,48±0,82
|
15,78±0,71*, ∆
|
|
Концентрация АТФ, мкмоль/л
|
0,79±0,09
|
0,28 ±0,05*
|
0,47±0,04*, ∆
|
Примечание:
* – различия по отношению к группе I, р≤0,05; ∆ – различия между группами
после криоконсервации (группа II и группа III).
Сдерживающим
фактором окислительных процессов является усиление активности антиоксидантных
ферментов. В ходе исследования выявлено увеличение активности СОД и каталазы в
сперматозоидах после добавления H2 в среду для криоконсервации
(группа III) (табл. 3). Активность СОД и каталазы увеличилась на 42% и 74% соответственно
(р≤0,05) относительно нативных сперматозоидов
(группа I).
Изучение энергетического статуса сперматозоидов, показало, что после
криоконсервации концентрация АТФ в сперматозоидах была ниже чем в нативных
сперматозоидах (группа I) на 65%. При использовании H2
в ходе криоконсервации (группа III) содержание АТФ в
сперматозоидах повышалось на 67% относительно группы II.
Была
выявлена корреляционная зависимость между показателями метаболизма и фазовыми
характеристиками сперматозоидов при криоконсервации и использовании H2
на фоне криоконсервации (табл. 3). Анализ корреляционных зависимостей между фазовой
высотой и показателем окислительного метаболизма выявил тесную отрицательную корреляцию
во всех группах (R=-0,88 – I группа, R=-0,85 – II группа, R=-0,93 – III группа) и сильную корреляцию между фазовой
высотой и показателем энергетического метаболизма (R=0,88 – I группа, R=0,85 – II группа,
R=0,93 – III группа).
Таблица 3. Корреляционный анализ фазовой высоты и показателей метаболизма у различных групп
|
|
Фазовая высота/МДА
|
Фазовая высота/АТФ
|
|
Нативные сперматозоиды,
(группа I)
|
-0,88
|
0,84
|
|
Сперматозоиды после
криоконсервации, (группа II)
|
-0,85
|
0,86
|
|
Сперматозоиды после воздействия
молекулярным водородом и криоконсервации, (группа III)
|
-0,93
|
0,91
|
Корреляционные связи доказывают, что анализ фазовый высоты сперматозоидов полученный методом интерференционной микроскопии позволяет оценить совокупную метаболическую активность сперматозоидов и обладает наибольшей чувствительностью по сравнению с биохимическими методами.
Анализируя полученные результаты, следует учитывать, что при криоконсервации липиды и белки в жидком состоянии затвердевают, превращаясь в гель, образуя жёсткую и хрупкую структуру, более чувствительную к повреждениям [24]. Наше исследование показывает, что во время криоконсервации происходит потеря целостности как акросомной, так и плазматической мембран. Во время замораживания в сперматозоидах наблюдаются значительные метаболические изменения и сильно ухудшается функция митохондрий [25]. Биоэнергетическая функция митохондрий играет важную роль в сперматозоидах особенно для капитации, гиперактивации и акросомной реакции [26]. Полученные результаты демонстрируют, что использование H2 эффективно для защиты метаболизма сперматозоидов в процессе криоконсервации, вероятно, за счет способности стимулировать синтез энергии, подавляя избыточное образование активных форм кислорода. Эти свойства H2 могут обеспечить сохранение функциональных параметров сперматозоидов.
В ходе проведенного исследования, показано, что использование H2 при криоконсервации сохраняло количество подвижных сперматозоидов, количество быстрых сперматозоидов, а так же среднюю скорость сперматозоидов на уровне нативные клеток (табл. 4). Тогда как после криоконсервации сперматозоидов данные показатели были снижены.
Таблица 4. Влияние молекулярного водорода на показатели фертильности сперматозоидов
|
Критерии фертильности сперматозоидов
|
Нативные разбавленные сперматозоиды, (группа I)
|
Сперматозоиды после
криоконсервации, (группа II)
|
Сперматозоиды после
воздействия молекулярным водородом и криоконсервации, (группа III)
|
|
Подвижность, %
|
82,51±3,95
|
71,15±3,34*
|
79,62±3,60 ∆
|
|
Количество
подвижных, млн/доза
|
35,76±2,17
|
27,35±2,16*
|
33,71±2,03 ∆
|
|
|
Количество
быстрых, млн/доза
|
65,54±7,14
|
51,77±6,13*
|
58,98±6,55 ∆
|
|
Средняя
скорость, мкм/сек
|
85,62±3,54
|
74,53±2,48*
|
81,56±3,52 ∆
|
Примечание:
* –различия по отношению к группе I,
р≤0,05; ∆ – различия между группами после криоконсервации (группа II и группа III).
Учитывая, что скорость движения сперматозоидов является одним из наиболее информативных показателей качества спермы [27], выявленное увеличение количество подвижных, быстрых сперматозоидов и средней скорости сперматозоидов при действии H2 по сравнению с данными показателями при криоконсервации доказывает эффективность использования H2 в качестве криопротектора.
1. В ходе исследования доказана эффективность использования Н2 как новой стратегии защиты сперматозоидов при криоконсервации.
2. Анализ интерфореграмм сперматозоидов дает комплексную оценку cоcтояния метаболических процессов сперматозоидов в ходе криоконсервации.
3. Фазовые изображения позволяют четко определить сперматозоиды с пониженным функциональным состоянием, что можно использовать для экспресс-анализа качества сперматозоидов.
Работа выполнена при поддержке Российского научного фонда (проект № 23-26-00205).
1. Watson P.F. The causes of reduced fertility with cryopreserved semen // Anim Reprod Sci. 2000. № 60–61. Р. 481–92. doi: 10.1016/s0378-4320(00)00099-3.
2. Sion B., Janny L., Boucher D., Grizard G. Annexin V binding to plasma membrane predicts the quality of human cryopreserved spermatozoa // Int J Androl. 2004. №27(2). Р. 108–1014. doi: 10.1046/j.1365-2605.2003.00457.x.
3. Kogan T., Dahan D.G., Laor R., Argov-Argaman N., Komsky-Elbaz A., Kalo D., Roth Z. Association between Fatty Acid Composition, Cryotolerance and Fertility Competence of Progressively Motile Bovine Spermatozoa // Animals (Basel). 2021. №11(10). Р. 2948. doi: 10.3390/ani11102948.
4. Thomson L.K., Fleming S.D., Aitken R.J., De Iuliis G.N., Zieschang J.A., Clark A.M. Cryopreservation-induced human sperm DNA damage is predominantly mediated by oxidative stress rather than apoptosis // Hum Reprod. 2009. № 24(9). Р. 2061–70. doi: 10.1093/humrep/dep214
5. Partyka A., Lukaszewicz E., Nizanski W., Twardon J. Detection of lipid peroxidation in frozen-thawed avian spermatozoa using C(11)-BODIPY(581/591) // Theriogenology. 2011. №75(9). Р. 1623–9. doi: 10.1016/j.theriogenology.2011.01.002.
6. Zhai X., Chen X., Shi J., Shi D., Ye Z., Liu W. Lactulose ameliorates cerebral ischemia-reperfusion injury in rats by inducing hydrogen by activating Nrf2 expression // Free Radic. Biol. Med. 2013. № 65. Р. 731–741. doi: 10.1016/j.freeradbiomed.2013.08.004
7. Miller M. W., Sadeh N. Traumatic stress, oxidative stress and post-traumatic stress disorder: neurodegeneration and the accelerated-aging hypothesis // Mol. Psychiatry 2014. № 19. Р. 1156–1162. doi: 10.1038/mp.2014.111
8. Lue N., Popescu G., Ikeda T., Dasari R., Badizadegan K., Feld M. Live cell refractometry using microfluidic devices // Opt. Lett. 2006. № 31(18). Р. 2759-2761. doi: 10.1364/ol.31.002759
9. Park Y., Popescu G., Badizadegan K., Dasari R., Feld M. Diffraction phase and fluorescence microscopy // Opt. Expr. 2006. № 14(18). Р. 8263-8268. doi: 10.1364/oe.14.008263
10. Левин Г.Г., Булыгин Ф.В., Вишняков Г.Н. Когерентные осцилляции состояния молекул белка в живых клетках // Цитология. 2005. Т.47. №4. С.348-356.
11. Дерюгина А.В., Иващенко М.Н., Игнатьев П.С., Метелин В.Б., Таламанова М.Н. Возможности прижизненной визуализации клеток крови при стрессе для оценки состояния организма // Научная визуализация. 2023. Т. 15. № 1. С. 90-99. doi: 10.26583/sv.15.1.08
12. Тычинский В.П., Кретушев А.В., Клемяшов И.В., Вышенская Т.В., Штиль А.А., Зацепина О.В. Когерентная фазовая микроскопия – новый подход к исследованию физиологического состояния ядрышка // ДАН. 2005. № 405(4). С. 432-436.
13. Deryugina A.V., Danilova D.A., Pichugin V.V., Brichkin Yu.D. The effect of molecular hydrogen on functional states of erythrocytes in rats with simulated chronic heart failure // Life. 2023. №3(13). Р. 418. doi: 10.3390/life13020418
14. Сирота Т.В. Стандартизация и регуляция скорости супероксидгенерирующей реакции автоокисления адреналина, используемой для определения про/антиоксидантных свойств различных материалов // Биомедицинская химия. 2016. № 6. С. 650–655. doi: 10.18097/PBMC20166206650
15. Дерюгина А.В., Абаева О.П., Романов С.В., Ведунова М.В., Рябова Е.Н., Васенин С.А., Титова Н.А. Электрокинетические, оксидантные и агрегационные свойства эритроцитов в послеоперационном периоде при трансплантации почки // Вестник трансплантологии и искусственных органов. 2020. № 2. С. 72-79. doi: 10.15825/1995-1191-2020-2-72-79
16. Дерюгина А.В., Бояринов Г.А., Симутис И.С., Никольский В.О., Кузнецов А.Б., Ефимова Т.С. Коррекция озонированной эритроцитной массой метаболических показателей эритроцитов и структуры миокарда после острой кровопотери // Цитология. 2018. № 2 (60). С. 89-95. doi: 10.31116/tsitol.2018.02.03
17. Дерюгина А.В., Иващенко М.Н., Белов А.А., Игнатьев П.С., Метелин В.Б. Применение лазерной интерференционной микроскопии для оценки функционального состояния эритроцитов // Цитология. 2021. № 1. С. 74-79. doi: 10.31857/S004137712101003X
18. Bailey J.L., Bilodeau J.F., Cormier N. Semen cryopreservation in domestic animals: a damaging and capacitating phenomenon // J. Androl. 2000. № 21. Р. 1-7.
19. Тычинский В. П. Динамическая фазовая микроскопия: возможен ли “диалог” с клеткой? // УФН. 2007. №5 (177). С. 535–552 doi: https://doi.org/10.3367/UFNr.0177.200705c.0535
20. Kondoh G., Tojo H., Nakatani Y., Komazawa N., Murata C., Yamagata K., Maeda Y., Kinoshita T., Okabe M., Taguchi R. Takeda J. Angiotensin-Converting Enzyme Is a GPI-Anchored Protein Releasing Factor Crucial for Fertilization // Nature Medicine. 2005. № 11. Р. 160-166. https://doi.org/10.1038/nm1179.
21. Юcипович А.И., Беpеcтовcкая Ю.Ю., Шутова В.В., Левин Г.Г., Геpаcименко Л.М., Макcимов Г.В., Pубин А.Б. Новые возможноcти иccледования микpобиологичеcкиx объектов методом лазеpной интеpфеpенционной микpоcкопии // Биофизика. 2011. Т. 56. Вып. 6. С. 1091–1098.
22. Загубиженко М.В., Юсипович А.И., Пирутин С.К., Минаев В.Л., Кудряшов Ю.Б. Использование метода лазерной интерференционной микроскопии для исследования состояния перитонеальных макрофагов мыши, облученных ультрафиолетовым светом // Радиационная биология. Радиоэкология. 2011. № 6(51). С. 715–720
23. Браже А.Р., Браже Н.А., Сосновцева О.В., Павлов А.Н., Мозекильде Э., Максимов Г.В. Исследование клеточной динамики с помощью интерференционной микроскопии с применением вейвлет-анализа // Компьютерные исследования и моделирование. 2009. Т. 1. № 1. С. 77–83
24. Gao D., Critser J.K. Mechanisms of cryoinjury in living cells // ILAR J. 2000. №41(4). Р. 187–96. doi: 10.1093/ilar.41.4.187.
25. Schober D., Aurich C., Nohl H., Gille L. Influence of cryopreservation on mitochondrial functions in equine spermatozoa // Theriogenology. 2007. № 68. Р. 745–54. doi: 10.1093/ilar.41.4.187.
26. O’Flaherty C., de Lamirande E., Gagnon C.. Positive role of reactive oxygen species in mammalian sperm capacitation: triggering and modulation of phosphorylation events // Free Radic Biol Med. 2006. № 41(4). Р. 528–40. doi: 10.1016/j.freeradbiomed.2006.04.027
27. Ляшенко А.А. Биологические показатели спермы быков в зависимости от срока хранения в жидком азоте // Зоотехническая наука Беларуси. 2015. Т. 50. №1. С. 126-134.
Using Sperm Imaging with Laser Interference Microscopy for Comprehensive Assessment of the Functional State of Cells during Cryopreservation and under the Action of Molecular Hydrogen
Authors: A.V. Deryugina1,A, M.N. Ivaschenko2,A,B, P.S. Ignatiev3,C, V.B. Metelin4,D
A Lobachevsky Nizhny Novgorod State University, Nizhny Novgorod, Russia
B Nizhny Novgorod State Agrotechnological University named after L.Ya. Florentyev, Nizhny Novgorod, Russia
C Production Association «Ural Optical-Mechanical Plant named after E.S. Yalamov», Yekaterinburg. Russia
D Moscow Regional Research Clinical Institute named after M.F. Vladimirsky, Moscow, Russia
1 ORCID: 0000-0001-8812-8559, derugina69@yandex.ru
2 ORCID: 0000-0001-6642-8518, marina.31@rambler.ru
3 ORCID: 0000-0001-5075-7034, ignasha2000@yandex.ru
4 ORCID: 0000-0003-0600-5729, verrv01@gmail.com
Abstract
Significant advances have been made in sperm cryopreservation but the search for effective sperm cryopreservation technologies is a pressing issue in modern biology and medicine. The most effective cryopreservation leaves 50-60% of viable cells. The paper discusses the use of molecular hydrogen (H2) as a new approach to enhancing sperm protection during freezing and thawing. H2 is a universal antioxidant and limits damage to biomolecules. Visual registration of spermatozoa under the action of H2 was performed using modern microscopy techniques. Laser interference microscopy was used in the work. Laser interference microscopy records the cell surface architectonics depending on the modulation of the optical density of cellular structures. This visualization option provides information on the metabolic level and expands the possibilities for interpreting experimental results. Sample preparation, dyes, and fixatives are not used in interference visualization. The paper presents an analysis of phase images of spermatozoa during cryopreservation and using H2 as a cryoprotector. Verification of the method for analyzing phase characteristics of spermatozoa as an indicator of the metabolic state of cells was performed by analyzing clinical and laboratory parameters of spermatozoa. The phase height of spermatozoa during cryopreservation decreased, the intensity of energy processes decreased, and the oxidative potential of cells increased. A direct correlation was shown between the phase height of spermatozoa and the concentration of ATP, and an inverse correlation was found from the concentration of malondialdehyde (MDA). The use of H2 determined an increase in the phase height of spermatozoa, an increase in energy metabolism, and a decrease in cell oxidation. Changes in the metabolic activity of spermatozoa under the action of H2 were combined with an improvement in sperm fertility. Thus, phase interference microscopy allows for a qualitative and quantitative assessment of the physiological state of spermatozoa. It is an objective method of vital analysis of complex metabolic activity of cells. It can be used for express diagnostics of their functional state.
Keywords: phase interference microscopy, diagnostic, sperm, molecular hydrogen.
1. Watson P.F. The causes of reduced fertility with cryopreserved semen // Anim Reprod Sci. 2000. № 60–61. Р. 481–92. doi: 10.1016/s0378-4320(00)00099-3.
2. Sion B., Janny L., Boucher D., Grizard G. Annexin V binding to plasma membrane predicts the quality of human cryopreserved spermatozoa // Int J Androl. 2004. №27 (2). Р. 108–1014. doi: 10.1046/j.1365-2605.2003.00457.x.
3. Kogan T., Dahan D.G., Laor R., Argov-Argaman N., Komsky-Elbaz A., Kalo D., Roth Z. Association between Fatty Acid Composition, Cryotolerance and Fertility Competence of Progressively Motile Bovine Spermatozoa // Animals (Basel). 2021. №11 (10). Р. 2948. doi: 10.3390/ani11102948.
4. Thomson L.K., Fleming S.D., Aitken R.J., De Iuliis G.N., Zieschang J.A., Clark A.M. Cryopreservation-induced human sperm DNA damage is predominantly mediated by oxidative stress rather than apoptosis // Hum Reprod. 2009. № 24(9). Р. 2061–70. doi: 10.1093/humrep/dep214
5. Partyka A., Lukaszewicz E., Nizanski W., Twardon J. Detection of lipid peroxidation in frozen-thawed avian spermatozoa using C(11)-BODIPY(581/591) // Theriogenology. 2011. №75 (9). Р. 1623–9. doi: 10.1016/j.theriogenology.2011.01.002.
6. Zhai X., Chen X., Shi J., Shi D., Ye Z., Liu W. Lactulose ameliorates cerebral ischemia-reperfusion injury in rats by inducing hydrogen by activating Nrf2 expression // Free Radic. Biol. Med. 2013. № 65. Р. 731–741. doi: 10.1016/j.freeradbiomed.2013.08.004
7. Miller M. W., Sadeh N. Traumatic stress, oxidative stress and post-traumatic stress disorder: neurodegeneration and the accelerated-aging hypothesis // Mol. Psychiatry 2014. № 19. Р. 1156–1162. doi: 10.1038/mp.2014.111
8. Lue N., Popescu G., Ikeda T., Dasari R., Badizadegan K., Feld M. Live cell refractometry using microfluidic devices // Opt. Lett. 2006. № 31(18). Р. 2759-2761. doi: 10.1364/ol.31.002759
9. Park Y., Popescu G., Badizadegan K., Dasari R., Feld M. Diffraction phase and fluorescence microscopy // Opt. Expr. 2006. № 14(18). Р. 8263-8268. doi: 10.1364/oe.14.008263
10. Levin G.G., Bulygin F.V., Vishnyakov G.N. Coherent oscillations of the state of protein molecules in living cells // Tsitology. 2005. V.47. №4. P.348-356.
11. Deryugina A.V., Ivaschenko M.N., Ignatiev P.S., Metelin V.B., Talamanova M.N. Possibilities of intravital visualization of blood cells under stress to assess the state of the body // Scientific visualization.2023. V. 15. № 1. P. 90-99. doi: 10.26583/sv.15.1.08
12. Tychinsky V.P., Kretushev A.V., Klemyashov I.V., Vyshenskaya T.V., Shtil A.A., Zatsepina O.V. Coherent phase microscopy – a new approach to studying the physiological state of the nucleolus // DAN. 2005. № 405(4). P. 432-436.
13. Deryugina A.V., Danilova D.A., Pichugin V.V., Brichkin Yu.D. The effect of molecular hydrogen on functional states of erythrocytes in rats with simulated chronic heart failure // Life. 2023. №3(13). Р. 418. doi: 10.3390/life13020418
14. Sirota T.V. Standardization and regulation of the rate of superoxide-generating reaction of adrenaline autoxidation used to determine the pro/antioxidant properties of various materials // Biomedical Chemistry. 2016. № 6. P. 650–655. doi: 10.18097/PBMC20166206650
15. Deryugina A.V., Abaeva O.P., Romanov S.V., Vedunova M.V., Ryabova E.N., Vasenin S.A., Titova N.A. Electrokinetic, oxidative and aggregation properties of red blood cells in the postoperative period following kidney transplantation // Russian journal of transplantology and artificial organs. 2020. № 2. P. 72-79. doi: 10.15825/1995-1191-2020-2-72-79
16. Deryugina A.V., Efimova T.S., Boyarinov G.A., Nikolskiy V.O., Kuznetsov A.B., Simutis I.S. Correction of metabolic indicators of erythrocytes and myocardium structure with ozonized red blood-cell mass // Cell and Tissue Biology. 2018. V. 12. № 3. P. 207-212. doi: 10.1134/S1990519X18030033
17. Deryugina A.V., Belov A.A., Ivashchenko M.N., Ignatiev P.S., Metelin V.B. Assessing the functional state of red blood cells by using the laser interference microscopy // Cell and Tissue Biology. 2021. V. 15. № 4. P. 388-392. doi: 10.1134/S1990519X21040027
18. Bailey J.L., Bilodeau J.F., Cormier N. Semen cryopreservation in domestic animals: a damaging and capacitating phenomenon // J. Androl. 2000. № 21. Р. 1-7.
19. 19. Tychinsky V. P. Dynamic phase microscopy: is a “dialogue” with a cell possible? // Uspekhi Fizicheskikh Nauk. 2007. №5 (177). P. 535–552 doi: 10.3367/UFNr.0177.200705c.0535
20. Kondoh G., Tojo H., Nakatani Y., Komazawa N., Murata C., Yamagata K., Maeda Y., Kinoshita T., Okabe M., Taguchi R. Takeda J. Angiotensin-Converting Enzyme Is a GPI-Anchored Protein Releasing Factor Crucial for Fertilization // Nature Medicine. 2005. № 11. Р. 160-166. https://doi.org/10.1038/nm1179.
21. Yusipovich A.I., Berestovskaya Yu.Yu., Shutova V.V., Levin G.G., Gerasimenko L.M., Maksimov G.V., Rubin A.B. New possibilities of studying microbiological objects by laser interference microscopy // Biophysics. 2011. V. 56. № 6. P. 1091–1098.
22. Zagubizhenko M.V., Yusipovich A.I., Pirutin S.K., Minaev V.L., Kudryashov Yu.B. Using the method of laser interference microscopy to study the state of mouse peritoneal macrophages irradiated with ultraviolet light // Radiation biology. Radioecology. 2011. № 6(51). P. 715–720
23. Brazhe A.R., Brazhe N.A., Sosnovtseva O.V., Pavlov A.N., Mozekilde E., Maksimov G.V. Study of cellular dynamics using interference microscopy with wavelet analysis // Computer research and modeling. 2009. V. 1. № 1. P. 77–83
24. Gao D., Critser J.K. Mechanisms of cryoinjury in living cells // ILAR J. 2000. №41(4). Р. 187–96. doi: 10.1093/ilar.41.4.187.
25. Schober D., Aurich C., Nohl H., Gille L. Influence of cryopreservation on mitochondrial functions in equine spermatozoa // Theriogenology. 2007. № 68. Р. 745–54. doi: 10.1093/ilar.41.4.187.
26. O’Flaherty C., de Lamirande E., Gagnon C.. Positive role of reactive oxygen species in mammalian sperm capacitation: triggering and modulation of phosphorylation events // Free Radic Biol Med. 2006. № 41(4). Р. 528–40. doi: 10.1016/j.freeradbiomed.2006.04.027
27. Lyashenko A.A. Biological parameters of bull sperm depending on the storage period in liquid nitrogen // Zootechnical science of Belarus. 2015. V. 50. №1. С. 126-134.